Fett er en viktig energikilde, og i artikkelen om betaoksidasjon så vi på hvordan fettsyrene brytes ned for å frigi denne energien. Men fett er også vår viktigste lagringsform for energi, og vi kan syntetisere fettsyrer for å lagre energioverskudd til senere bruk. I denne artikkelen tar vi for oss hvordan vi syntetiserer fettsyrer, og til slutt litt om hvordan nebrytning og syntese av fettsyrer reguleres.
Denne artikkelen tilhører artikkelserien om metabolismen.
Relevante artikler:
Fettsyresyntesen er en reversering av reaksjonene vi så ved β-oksidasjonen. Selv om prosessene er motsatte av hverandre, er det også et par ting som skiller dem.
- Mens nedbrytningen skjer inni mitokondriene (og peroksisomene), foregår syntesen i cellens cytosol.
- Der den krympende fettsyren er bundet til Koenzym A (CoA), er den voksende fettsyren bundet til acylbærende protein (ACP).
- Enzymene som er involvert i fettsyresyntesen er koblet sammen i et stort enzymkompleks som kalles fettsyre syntase, mens enzymene i betaoksidasjonen fungerer som enkeltstående enzymer. Fettsyre syntase elongerer fettsyren til den består av 16 karbonatomer.
- Β-oksidasjonen fører til en frigjøring av tokarbonenheten acetyl-CoA, mens fettsyresyntesen fester på to karboner fra trekarbonenheten malonyl-ACP.
Når vi produserer nye fettsyrer så kalles prosessen ofte de novo lipogenese, som betyr nydanning av fettsyrer, men her brukes bare fettsyresyntese.
Stegene i fettsyresyntesen
Utgangspunktet for fettsyresyntesen er acetyl-ACP, og trekarbonenheten malonyl-ACP donerer to karbonatomer til den voksende fettsyren gjennom en prosess som katalyseres av fettsyre syntase. Det hastighetsregulerende steget i fettsyresyntesen er produksjonen av Malonyl-CoA, som dannes fra acetyl-CoA i en irreversibel prosess ved hjelp av enzymet acetyl-CoA karboksylase, som bruker biotin som kofaktor. Produksjonen av malonyl-CoA er energikrevende, og prosessen koster et ATP-molekyl.
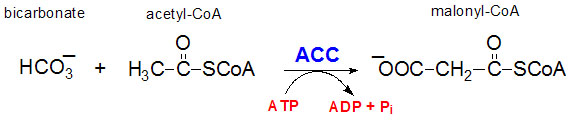
Bilde: www.themedicalbiochemistrypage.org
ACP er en polypeptidkjede, og binder til andre molekyler omtrent på samme måte som CoA. Acetyl-CoA og malonyl-CoA omdannes til acetyl-ACP og malonyl-ACP ved hjelp av enzymene acetyl- og malonyl transacylase. Som nevnt er det malonyl-ACP som donerer to karboner til den voksende fettsyren, og utgangspunktet for produksjonen av en ny fettsyre er acetyl-ACP.
Fettsyre syntase er enzymkomplekset som katalyserer koblingen av tokarbongruppen fra malonyl-CoA, og dette foregår i fire steg. Dette er en reversering av det som foregikk under β-oksidasjonen.
- Det første steget er en kondensering/sammenslåing av Acetyl-ACP og Malonyl-ACP, og produktet av dette er acetoacetyl-ACP, som er et ketoacyl-ACP med fire karbonatomer i kjeden. I denne reaksjonen frigjøres et karbonmolekyl fra malonyl-ACP i form av karbondioksid, og ACP fra acetyl-ACP frigjøres også for å gjøre plass til de nye karbonatomene. Reaksjonen katalyseres av acyl-malonyl-ACP kondenseringsenzym. Dette steget er det motsatte av steg 4 i β-oksidasjonen, der acetyl-CoA ble spaltet av.
- Det andre steget er en reduksjon der det dannes et hydroksyacyl-ACP. Denne reduksjonen skjer ved hjelp av enzymet β-ketoacyl ACP reduktase. Reduksjonsagenten er NADPH, som donerer et hydrogenatom og oksideres til NADP+. Denne reaksjonen er det motsatte av steg 3 i β-oksidasjonen, som var en oksideringsreaksjon.
- Det tredje steget er en dehydreringsreaksjon, der vi ved å fjerne et vannmolekyl får dannet en enoyl-ACP. Reaksjonen drives av enzymet 3-hydroksyacyl ACP dehydratase. Dette steget er det motsatte av det andre steget i β-oksidasjonen, der enoyl-CoA fikk koblet på et vannmolekyl og dannet hydroksyacyl-CoA.
- Det siste steget i fettsyresyntesen er en ny reduseringsreaksjon, der sluttproduktet er en fettsyre som er to karbonatomer lenger enn utgangspunktet. Denne reaksjonen drives av enzymet enoyl ACP reduktase, og igjen er NADPH reduksjonsagent. Dette er det motsatte av den første reaksjonen i β-oksidasjonen, der vi fikk dannet en enoyl-CoA.
For hver runde av fettsyresyntesen vokser altså fettsyren med to karbonatomer. Sluttproduktet for fettsyresyntesen er en mettet fettsyre med 16 karbonatomer, palmitinsyre. Enzymet thioesterase spalter av ACP slik at vi sitter igjen med en ferdig fettsyre.
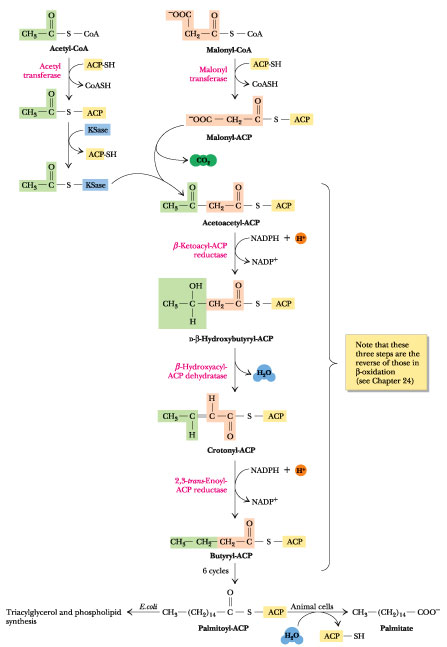
Alle karbonatomene i den endelige fettsyren kommer fra acetyl-CoA (som først omdannes til malonyl-CoA), som vi kan få fra nedbrytning av fettsyrer eller glukose. Når vi lagrer overskuddsenergi fra glukose som fett så må glukosen først gå gjennom glykolysen for å danne acetyl-CoA, som deretter går inn i fettsyresyntesen for å produsere fettsyrer. Produksjonen av acetyl-CoA skjer i mitokondriene, og acetyl-CoA må kobles til citrat (intermediat i sitronsyresyklusen) for å fraktes ut i cytosol der fettsyresyntesen foregår.
Vi har egne elongeringsenzymer som gjør at vi kan forlenge fettsyrene og dermed produsere lengre fettsyrer enn palmitinsyre. Vi har også egne desaturaser som kan sette inn dobbeltbindinger i karbonkjeden slik at vi får dannet umettede fettsyrer. Disse prosessene forklares i en egen artikkel.
Kontroll av fettsyremetabolismen
Syntese og nedbrytning av fett reguleres for å passe til kroppens fysiologiske behov. Når cellene har god tilgang på energi og karbohydrater så prioriteres fettsyresyntese, mens β-oksidasjonen prioriteres i motsatt tilfelle. Det viktigste kontrollpunktet er enzymet Acetyl-CoA karboksylase, som er det hastighetsregulerende enzymet i fettsyresyntesen ved å produsere malonyl-CoA i en irreversibel reaksjon. Malonyl-CoA hindrer transport av fettsyrer inn i mitokondriet ved å effektivt hemme carnitin palmitoyl acyltransferase 1 som er enzymet som styrer denne transporten. Dette gjør at syntese og nedbrytning av fettsyrer ikke skjer samtidig. Aktiviteten til acetyl-CoA karboksylase reguleres på flere måter.
Enzymet hemmes ved fosforylering, og aktiveres ved defosforylering. Fosforyleringen styres av cellens tilgang på energisubstrat, og når tilgangen på dette er høy så aktiveres enzymet slik at fettsyresyntesen settes i gang, mens fettsyrenedbrytning favoriseres når tilgangen på energi er lav.
Acetyl-CoA karboksylase reguleres også allosterisk ved binding av citrat. Mengden citrat øker når tilgangen på acetyl-CoA og ATP er høy inne i cellen, og signaliserer derfor et overskudd av energi. Da aktiveres enzymet. Palmitoyl-CoA er en enoyl-CoA vi har mye av når tilgangen på fettsyrer er stor. Dette hemmer karboksylasen slik at fettsyresyntesen bremses.
Hormonell regulering av fettmetabolismen skjer først og fremst ved hormonene insulin, glukagon og adrenalin. Overordnet stimulerer insulin fettsyresyntesen ved å aktivere acetyl-CoA karboksylase, mens glukagon og adrenalin har motsatt effekt. I tillegg virker disse hormonene inn på frigjøring og innlagring av fettsyrer i fettvev. Glukagon og adrenalin vil stimulere hormonsensitiv lipase (HSL) som sørger for frigjøring av fettsyrer fra triglyseridene i fettvevet. Insulin på sin side vil stimulere opptaket av fettsyrer fra blodet ved å stimulere lipoprotein lipase, samt hemme HSL og dermed frigjøring av fettsyrer fra fettvev.
Artikkelen er sist oppdatert august 2013
Tilbake til artikkelserien om metabolismen.
«Glukagon og adrenalin vil stimulere hormonsensitiv lipase (HSL) som sørger for frigjøring av fettsyrer fra triglyseridene i fettvevet. Insulin på sin side vil stimulere opptaket av fettsyrer fra blodet ved å stimulere lipoprotein lipase, samt hemme HSL og dermed frigjøring av fettsyrer fra fettvev.»
Jeg klarer ikke se forskjellen.. Begge frigjør fettsyrer fra fettvev?
Leser den om og om igjen. Min feil.. Det var bare en veldig innviklet setning..
Hei, så bra det ordnet seg! Jeg er enig i at den var litt komplisert.
Kan man se denne prosessen på molekylnivå noe sted på internet. Morsomt at glukose skal bli fett. Er det mange ernæringsfysiologer som tør å gå i mot de norske kostholdsrådene som sier mer karbohydrater og mindre fett.